But a phone could conceivably collect pollen and redistribute, no? WHAT AN ADDED FEATURE!!!!
I kid. What a fucking dystopia we don't know we're living in.
Moderators: Elvis, DrVolin, Jeff
![]() by 82_28 » Sat May 14, 2016 12:28 pm
by 82_28 » Sat May 14, 2016 12:28 pm

![]() by smoking since 1879 » Sat May 14, 2016 12:40 pm
by smoking since 1879 » Sat May 14, 2016 12:40 pm
82_28 » Sat May 14, 2016 5:28 pm wrote:Maybe an added feature in the phones that pollinates?
But a phone could conceivably collect pollen and redistribute, no? WHAT AN ADDED FEATURE!!!!
I kid. What a fucking dystopia we don't know we're living in.

![]() by Cordelia » Wed Jun 29, 2016 10:33 am
by Cordelia » Wed Jun 29, 2016 10:33 am


![]() by Burnt Hill » Thu Sep 01, 2016 7:01 pm
by Burnt Hill » Thu Sep 01, 2016 7:01 pm
‘Like it’s been nuked': Millions of bees dead after South Carolina sprays for Zika mosquitoes
On Sunday morning, the South Carolina honey bees began to die in massive numbers.
Death came suddenly to Dorchester County, S.C. Stressed insects tried to flee their nests, only to surrender in little clumps at hive entrances. The dead worker bees littering the farms suggested that colony collapse disorder was not the culprit — in that odd phenomenon, workers vanish as though raptured, leaving a living queen and young bees behind.
Instead, the dead heaps signaled the killer was less mysterious, but no less devastating. The pattern matched acute pesticide poisoning. By one estimate, at a single apiary — Flowertown Bee Farm and Supply in Summerville — 46 hives died on the spot, totaling some 2.5 million bees.
http://www.msn.com/en-us/news/us/%E2%80%98like-it%E2%80%99s-been-nuked-millions-of-bees-dead-after-south-carolina-sprays-for-zika-mosquitoes/ar-AAim4q3?li=BBmkt5R&ocid=spartanntp

![]() by Sounder » Thu Sep 01, 2016 7:33 pm
by Sounder » Thu Sep 01, 2016 7:33 pm

![]() by Harvey » Thu Oct 20, 2016 8:45 am
by Harvey » Thu Oct 20, 2016 8:45 am

![]() by Cordelia » Tue Oct 10, 2017 7:23 am
by Cordelia » Tue Oct 10, 2017 7:23 am
'Insane' discovery: 30,000 bees - and 40 pounds of honey - inside the walls of a New Jersey house
The honey bees had taken up residence inside the walls of the house.
A homeowner in Hillside, New Jersey, near Newark, told CBS New York it was "really noisy" with honey bees "humming and huddled up together."
But he had no idea just how many bees had been lurking behind those walls.
In fact, there were about 30,000 of them - and about 40 pounds of honey, according to a beekeeper who was called to remove them.
Video showed the moments after the beekeeper opened one of the walls last month.
"This is insane," Mickey Hegedus said in the video.
"What I'm doing is I'm slowly cutting out each piece of lath - and as I do it, it just exposes more bees and more honey and more comb," the third-generation beekeeper said in the video, as bees buzzed and darted from a large hole in the wall and latched onto his protective suit. "These are Africanized - these are the most aggressive bees I think I've ever cut out of a hive."
Hegedus, known in the New York and New Jersey area as Mickey the Beekeeper, told The Washington Post that it appeared that the bees had been getting into the house through an outside opening meant for electrical wiring.
Because these bees are so aggressive, they're better foragers and collect more honey and build more comb than most bees; they're also more defensive, he said. All of those factors, along with a warm spring, contributed to the hive's large size.
Hegedus was called to the home Sept. 30 to remove them.
Generally, exterminating bees is considered a last option, according to the New Jersey Beekeepers Association, which says honey bees "should be left alone unless their hive is in conflict with human activity."
In such cases, the bees can be removed but "should be killed only as a last resort. Except for colonies residing within buildings or other indoor structures, it is illegal in some states (e.g. NJ) to kill honey bee colonies without approval from appropriate agencies," according to an association pamphlet.
After Hegedus confirmed that they were honey bees, he said, he drilled a small hole in the wall and put in a camera to see the swarm behind it.1
One or two of the bees escaped from the small hole and Hegedus said, "one flew right at my head and into my hair and started stinging me."
When he opened the wall, he said, "they were all over me." He said he got stung about 30 times, mainly around his wrists where his gloves met his protective suit.
Hegedus said he used a special vacuum attached to a beehive box to gently suck some of the bees from the wall cavity - and off himself.
"I had to - they wouldn't stop," he said, adding that he then cut the comb and put the pieces into other beehive boxes - a process that he said took about five hours.
Hegedus said he was able to save about 95 percent of the hive, but because the bees were so aggressive, he was not able to keep them or give them away.
Instead, he said, he released them into an isolated area in Mountainside, New Jersey.
http://jacksonville.com/news/national/2 ... alls-house

![]() by seemslikeadream » Thu Nov 30, 2017 1:32 pm
by seemslikeadream » Thu Nov 30, 2017 1:32 pm
A Clue in the Bee Death Mystery
Insecticides are often blamed, but new signs point to another chemical.
Tom PhilpottNov. 29, 2017 6:00 AM
A rusty-patched bumble bee. Smithsonian's National Zoo/Flickr
Domesticated honeybees get all the buzz, but wild bumble bees are in decline too, both globally and here in the United States. What gives? It’s an important question, because while managed honeybees provide half of the pollination required by US crops, bumble and other wild bees deliver the other half.
Insecticides used in agriculture are one possible trigger—they exist to kill insects, after all, and bumble bees are insects. But a different kind of farm chemical, one designed to kill fungi that harm crops, has emerged as a possible culprit. A new study by a team of researchers led by Cornell University entomologist Scott McArt adds to the growing dossier of studies pinpointing fungicide as a potential bee killer (see here, here, and here).
In their paper, McArt and his team looked at two factors related to bumble bee decline. The first is that many bumble bee species appear to be confining themselves to ever smaller geographical regions—a phenomenon known as range contraction. The other is a microscopic parasite called Nosema bombi, which has turned up at high rates in bumble bee species known to be deteriorating.
Many bumble bee species appear to be confining themselves to ever smaller geographical regions.
The team analyzed samples of bumble bees from eight species taken by previous researchers from 284 sites across the country between 2007 and 2009. For the areas surrounding each site, the McArt team crunched data for 24 environmental factors that might affect bee health: Everything from the level of nearby residential development to the portion of land devoted to forests to the amount of insecticides, herbicides, and fungicides applied by farmers.
Their goal was to see which of these factors was most closely associated with shrinking habitats and Nosema bombi infections. Total fungicide applications in a given area emerged as the best predictor of range contraction; and a single widely used fungicide, chlorothalonil, proved to be the clearest indicator of Nosema bombi prevalence.
The result wasn’t a total surprise. A 2015 study by University of Wisconsin and US Department of Agriculture researchers found that bumble bee hives exposed to small amounts of chlorothalonil—which is widely used in fruits, vegetables, and orchard crops—”produced fewer workers, lower total bee biomass, and had lighter mother queens than control colonies.”
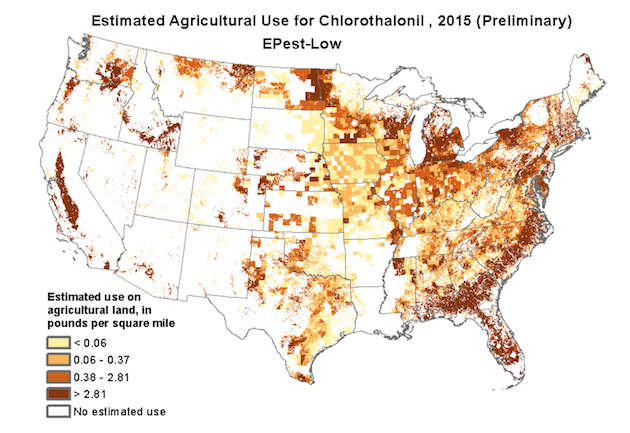
Here’s a map of where chlorothalonil is used, from the US Geological Survey:
To make matters worse, the fungicides can exacerbate the effect of other types of chemicals. “While most fungicides are relatively nontoxic to bees, many are known to interact synergistically with insecticides, greatly increasing their toxicity to the bees,” McArt said in a press release accompanying the study.
Meanwhile, bumblebees continue to struggle. In its final days, the Obama administration declared the rusty-patched bumble bee, pictured above, as endangered—and its plight is so severe that the Trump administration opted to secure the designation, after hinting it would undo it.
Unlike murders, ecological collapses rarely have a single perpetrator. Rather, they’re caused by a host of culprits working together in ways that are hard to unpack. Add fungicides to the list of potential offenders in the ongoing pollinator crisis.
http://www.motherjones.com/food/2017/11 ... h-mystery/

![]() by smoking since 1879 » Thu Nov 30, 2017 5:55 pm
by smoking since 1879 » Thu Nov 30, 2017 5:55 pm
![]() by seemslikeadream » Thu Nov 30, 2017 6:27 pm
by seemslikeadream » Thu Nov 30, 2017 6:27 pm
Tumbling bumblebee populations linked to fungicides -- ScienceDaily
Instead, they found a shocker: fungicides, commonly thought to have no impact.
"Insecticides work; they kill insects. Fungicides have been largely overlooked because they are not targeted for insects, but fungicides may not be quite as benign -- toward bumblebees -- as we once thought. This surprised us," said Scott McArt, assistant professor of entomology and the lead author on a new study, "Landscape Predictors of Pathogen Prevalence and Range Contractions in United States Bumblebees," published Nov. 15 in the journal Proceedings of the Royal Society B.
This new work shows how fungicides -- particularly chlorothalonil, a general-use fungicide often found in bumblebee and honeybee hives -- may negatively affect bee health, said McArt, a fellow at Cornell's Atkinson Center for a Sustainable Future.
Building on a large data set collected by Sydney Cameron, professor of entomology at the University of Illinois, the scientists discovered what they call "landscape-scale" connections between fungicide usage, pathogen prevalence and declines of endangered United States bumblebees.
While fungicides control plant pathogens in crops, the bees pick up their residue when foraging for pollen and nectar. As farms use both insecticides and fungicides, the scientists worry about synergy. "While most fungicides are relatively nontoxic to bees, many are known to interact synergistically with insecticides, greatly increasing their toxicity to the bees," McArt said.
Chlorothalonil has been linked to stunted colony growth in bumblebees and an increased vulnerability to Nosema, a fatal gut infection in bumblebees and honeybees.
"Nosema can be devastating to bumblebees and honeybees," said McArt. "Since fungicide exposure can increase susceptibility of bees to Nosema, this may be the reason we're seeing links between fungicide exposure, Nosema prevalence and bumblebee declines across the United States in this data set."
For domestic and global agriculture, bumblebees are a key component due to their ability to use "buzz pollination" that vibrates and shakes pollen loose from flowers. In the United States, bees contribute more than $15 billion to the economy and $170 billion to global agribusiness, according to global economic research and a 2012 Cornell study. While half of crop pollination work is done by commercially managed honeybees in the U.S., the other half is done by bumblebees and wild bees. In New York, pollination services contribute $500 million to the state's agricultural economy.
McArt and his Cornell colleagues will continue to investigate fungicide-insecticide synergisms and fungicide-pathogen interactions under the New York State Pollinator Protection Plan and a new grant from the New York Farm Viability Institute.
Funding was provided by the U.S. Department of Agriculture and the National Science Foundation.
Materials provided by Cornell University. Original written by Joe Schwartz. Note: Content may be edited for style and length.
https://www.sciencedaily.com/releases/2 ... 195041.htm
Disruption of quercetin metabolism by fungicide affects energy production in honey bees (Apis mellifera).
Mao W1, Schuler MA2, Berenbaum MR3.
Author information
Abstract
Cytochrome P450 monooxygenases (P450) in the honey bee, Apis mellifera, detoxify phytochemicals in honey and pollen. The flavonol quercetin is found ubiquitously and abundantly in pollen and frequently at lower concentrations in honey. Worker jelly consumed during the first 3 d of larval development typically contains flavonols at very low levels, however. RNA-Seq analysis of gene expression in neonates reared for three days on diets with and without quercetin revealed that, in addition to up-regulating multiple detoxifying P450 genes, quercetin is a negative transcriptional regulator of mitochondrion-related nuclear genes and genes encoding subunits of complexes I, III, IV, and V in the oxidative phosphorylation pathway. Thus, a consequence of inefficient metabolism of this phytochemical may be compromised energy production. Several P450s metabolize quercetin in adult workers. Docking in silico of 121 pesticide contaminants of American hives into the active pocket of CYP9Q1, a broadly substrate-specific P450 with high quercetin-metabolizing activity, identified six triazole fungicides, all fungal P450 inhibitors, that dock in the catalytic site. In adults fed combinations of quercetin and the triazole myclobutanil, the expression of five of six mitochondrion-related nuclear genes was down-regulated. Midgut metabolism assays verified that adult bees consuming quercetin with myclobutanil metabolized less quercetin and produced less thoracic ATP, the energy source for flight muscles. Although fungicides lack acute toxicity, they may influence bee health by interfering with quercetin detoxification, thereby compromising mitochondrial regeneration and ATP production. Thus, agricultural use of triazole fungicides may put bees at risk of being unable to extract sufficient energy from their natural food.
https://www.ncbi.nlm.nih.gov/pubmed/28193870
Colonies of Bumble Bees (Bombus impatiens) Produce Fewer Workers, Less Bee Biomass, and Have Smaller Mother Queens Following Fungicide Exposure.
Bernauer OM1, Gaines-Day HR2, Steffan SA3,4.
Author information
Abstract
Bees provide vital pollination services to the majority of flowering plants in both natural and agricultural systems. Unfortunately, both native and managed bee populations are experiencing declines, threatening the persistence of these plants and crops. Agricultural chemicals are one possible culprit contributing to bee declines. Even fungicides, generally considered safe for bees, have been shown to disrupt honey bee development and impair bumble bee behavior. Little is known, however, how fungicides may affect bumble bee colony growth. We conducted a controlled cage study to determine the effects of fungicide exposure on colonies of a native bumble bee species (Bombus impatiens). Colonies of B. impatiens were exposed to flowers treated with field-relevant levels of the fungicide chlorothalonil over the course of one month. Colony success was assessed by the number and biomass of larvae, pupae, and adult bumble bees. Bumble bee colonies exposed to fungicide produced fewer workers, lower total bee biomass, and had lighter mother queens than control colonies. Our results suggest that fungicides negatively affect the colony success of a native bumble bee species and that the use of fungicides during bloom has the potential to severely impact the success of native bumble bee populations foraging in agroecosystems.
https://www.ncbi.nlm.nih.gov/pubmed/26463198
Crop Pollination Exposes Honey Bees to Pesticides Which Alters Their Susceptibility to the Gut Pathogen Nosema ceranae
Jeffery S. Pettis,
Abstract
Recent declines in honey bee populations and increasing demand for insect-pollinated crops raise concerns about pollinator shortages. Pesticide exposure and pathogens may interact to have strong negative effects on managed honey bee colonies. Such findings are of great concern given the large numbers and high levels of pesticides found in honey bee colonies. Thus it is crucial to determine how field-relevant combinations and loads of pesticides affect bee health. We collected pollen from bee hives in seven major crops to determine 1) what types of pesticides bees are exposed to when rented for pollination of various crops and 2) how field-relevant pesticide blends affect bees’ susceptibility to the gut parasite Nosema ceranae. Our samples represent pollen collected by foragers for use by the colony, and do not necessarily indicate foragers’ roles as pollinators. In blueberry, cranberry, cucumber, pumpkin and watermelon bees collected pollen almost exclusively from weeds and wildflowers during our sampling. Thus more attention must be paid to how honey bees are exposed to pesticides outside of the field in which they are placed. We detected 35 different pesticides in the sampled pollen, and found high fungicide loads. The insecticides esfenvalerate and phosmet were at a concentration higher than their median lethal dose in at least one pollen sample. While fungicides are typically seen as fairly safe for honey bees, we found an increased probability of Nosema infection in bees that consumed pollen with a higher fungicide load. Our results highlight a need for research on sub-lethal effects of fungicides and other chemicals that bees placed in an agricultural setting are exposed to.
Figures
Citation: Pettis JS, Lichtenberg EM, Andree M, Stitzinger J, Rose R, vanEngelsdorp D (2013) Crop Pollination Exposes Honey Bees to Pesticides Which Alters Their Susceptibility to the Gut Pathogen Nosema ceranae. PLoS ONE 8(7): e70182. https://doi.org/10.1371/journal.pone.0070182
Editor: Fabio S. Nascimento, Universidade de São Paulo, Faculdade de Filosofia Ciências e Letras de Ribeirão Preto, Brazil
Received: March 25, 2013; Accepted: June 16, 2013; Published: July 24, 2013
This is an open-access article, free of all copyright, and may be freely reproduced, distributed, transmitted, modified, built upon, or otherwise used by anyone for any lawful purpose. The work is made available under the Creative Commons CC0 public domain dedication.
Funding: Funding for this study was provided by the National Honey Board (http://www.honey.com/) and the USDA-ARS Areawide Project on Bee Health (http://www.ars.usda.gov/research/projec ... _no=412796). Neither the Honey Board nor USDA-ARS Program Staff had a role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: Dennis vanEngesldorp is a PLOS ONE Editor. All other authors have declared that no competing interests exist. This does not alter the authors' adherence to all the PLOS ONE policies on sharing data and materials.
Introduction
Honey bees, Apis mellifera, are one of the most important pollinators of agricultural crops [1]. Recent declines in honey bee populations in many North American and European countries [2]–[4] and increasing cultivation of crops that require insects for pollination [5] raise concerns about pollinator shortages [5], [6]. Habitat destruction, pesticide use, pathogens and climate change are thought to have contributed to these losses [2], [7], [8]. Recent research suggests that honey bee diets, parasites, diseases and pesticides interact to have stronger negative effects on managed honey bee colonies [9], [10]. Nutritional limitation [11], [12] and exposure to sub-lethal doses of pesticides [13]–[16], in particular, may alter susceptibility to or severity of diverse bee parasites and pathogens.
Recent research is uncovering diverse sub-lethal effects of pesticides on bees. Insecticides and fungicides can alter insect and spider enzyme activity, development, oviposition behavior, offspring sex ratios, mobility, navigation and orientation, feeding behavior, learning and immune function [9], [13], [14], [16]–[22]. Reduced immune functioning is of particular interest because of recent disease-related declines of bees including honey bees [3], [23]. Pesticide and toxin exposure increases susceptibility to and mortality from diseases including the gut parasite Nosema spp. [14], [15]. These increases may be linked to insecticide-induced alterations to immune system pathways, which have been found for several insects, including honey bees [22], [24]–[26].
Surveys of colony food reserves and building materials (i.e. wax) have found high levels and diversity of chemicals in managed colonies [18], [27], [28]. These mixtures have strong potential to affect individual and colony immune functioning. However, almost all research to-date on pesticides’ effects on pathogen susceptibility fed a single chemical to test bees [16]. Because pesticides may have interactive effects on non-target organisms (e.g. [29]), it is crucial to determine how real world combinations and loads of pesticides affect bee health.
One pathogen of major concern to beekeepers is Nosema spp. The endoparasitic fungal infections of N. apis and N. ceranae adversely affect honey bee colony health, and can result in complete colony collapse [30]. Infection with Nosema in the autumn leads to poor overwintering and performance the following spring [31], and queens can be superseded soon after becoming infected with Nosema [32]. We chose Nosema as a model pathogen because earlier work [13], [14] had demonstrated an interaction with pesticide exposure.
This study addresses two important questions. 1) What types of pesticides might bees be exposed to in major crops? While multiple studies have characterized the pesticide profile of various materials inside a honey bee nest [27], [28], few have looked at the pollen being brought back to the nest. 2) How do field-relevant pesticides blends affect bees’ susceptibility to infection by the Nosema parasite?
Methods
Ethics Statement
Pollen was collected from honey bees with permission of the beekeepers and the land owners.
Hive Selection and Pollen Collection
We collected pollen carried by foraging honey bees returning to the hive for nine hives in seven crops: almond, apple, blueberry, cranberry, cucumber, pumpkin, and watermelon (Table 1). For each crop, we selected three fields that were separated by at least 3.2 km. Hives were deployed in these fields for pollination services based on growers’ needs. Within each selected field, we chose the three honey bee hives with the strongest foraging forces by observing flight in the bee yard for 5–10 min, and attached plastic pollen traps (Brushy Mountain Bee Farm, Moravian Falls, NC) to these hives. Pollen traps collect the pollen pellets bees carry on their hind tibiae in flattened regions called corbiculae. Bees use this pollen to make food for larvae inside the nest. We checked traps after three days, and removed them if they contained at least 5 g of pollen. Traps with less than 5 g remained on hives until they contained 5 g of pollen or for 10 days. We placed pollen removed from traps in 50 mL centrifuge tubes and stored the samples on ice until they could be transferred to a −29°C freezer in the lab.
thumbnail
Table 1. Quantity and diversity of pollen collected in pollen traps on individual honey bee hives.
https://doi.org/10.1371/journal.pone.0070182.t001
Because our first round of pollen trapping in cranberry fields yielded little pollen, we collected pollen from each hive in cranberry fields twice: early in the flowering season and late in the season. We separate these samples in data analyses, referring to them as “Cranberry early” and “Cranberry late.”
We measured the wet weight of each pollen sample, and compared the quantity of pollen collected by hives in different crops via a Kruskal-Wallis test followed by a post-hoc non-parametric Tukey-type test (using the R package nparcomp [33]). We then divided each sample into three portions. A 5 g subsample was sorted by color and then each group of similarly colored pollen pellets were identified (see below); a 3 g subsample was sent to the USDA’s Agricultural Marketing Service Laboratory in Gastonia, NC for pesticide analysis; and a 10 g subsample was sent to the USDA-ARS Bee Research Laboratory (Beltsville, MD) for the Nosema infection study. Because almond pollen was collected after all other pollens, we were unable to include it in the pesticide analysis and Nosema infection study. In cases where the total amount of pollen collected from a single colony was less than 6 g all the pollen was used for pesticide analysis.
Pollen Identification
Each 5 g pollen subsample was dehydrated in a drying oven at 40°C. We considered a sample to be dry when its weight did not change between two consecutive time points (measured every 4–6 h). Typically pollen dried in 12–18 h. To identify pollen types collected by the bees, we sorted the pollen in each subsample by color, quantified each color by comparing to Sherwin-Williams® color palettes, re-weighed after color separation and fixed each color from each subsample on a separate slide. We prepared each slide by grinding 2 pollen pellets in 2 mL water and letting them dissolve to form a slurry. We placed a small amount of slurry on a slide with a drop of silicon oil, and covered slides and sealed with clear nail polish after letting air bubbles escape for 48 h. We visually identified each pollen type under 400x magnification by comparing with published reference collections [34]–[36]. Visual identification of pollen grains through comparison with voucher or reference specimens is standard in pollination ecology [37], [38]. Similarities between closely related pollens, however, sometimes prevent identification to genus or species with this method [39]. Because of this limitation, we assumed that all pollen collected in apple (Malus domestica) orchards that was identified as Malus sp. was from apple trees, and that all pollen in the Cucurbitaceae family collected in cucumber (Cucurbitaceae, Cucumis sativus) fields was from cucumber flowers.
For each subsample, we estimated pollen diversity as the number of different pollen colors collected from that bee hive. We also calculated the proportion, by weight, of the pollen that was identified as belonging to the target crop’s genus. Many samples could only be identified to genus, so assessing target genus rather than target crop permitted a more inclusive analysis. We used Kruskal-Wallis tests to determine whether either of these measures differed with the crop in which sampled bee hives were placed.
Pesticide Analysis
We determined the identity and load of pesticide residues present in pollen samples collected from all crops (except almond). For each field sampled (n = 19), we pooled pollen from the three hives for analysis. One early-season cranberry field and one cucumber field did not yield sufficient pollen in traps for pesticide analysis. Methods follow the LC/MS-MS and GC/MS methods for pollen analysis described in Mullin et al. [27]. We used these data to determine the total number of pesticides detected in each sample, each sample’s total pesticide load, and the diversity and load of pesticides in each of 10 categories: insecticides, fungicides, herbicides, and several insecticide types (carbamates, cyclodienes, formamidines, neonicotinoids, organophosphates, oxadiazines and pyrethroids). To permit comparison between categories with different numbers of elements, we calculated diversity as the proportion of pesticides from a category found in a given sample, and load as the total load divided by the number of chemicals in that category. We only calculated diversity for categories with at least three chemicals.
The total number of pesticides present and total load did not meet parametric assumptions. We thus analyzed how these variables differ between crops using non-parametric Kruskal-Wallis tests. When separated by category and log-transformed, pesticide loads did meet parametric assumptions. We thus determined whether load varied by pesticide category using a general linear mixed model with sample as a random effect, to control for the fact that our regression included one data point per category from each sample. Insufficient degrees of freedom prevented us from expanding this model to include crop. We thus asked whether the pesticide load and diversity varied with crop for each category using one Kruskal-Wallis test per category and applying a sequential Bonferroni correction [40] across pesticide categories to control for multiple comparisons.
Nosema Infection
The Nosema infection experiment is similar to published methods [26]. We obtained 210 disease-free honey bees from each of three healthy colonies at the Bee Research Laboratory. Each bee was placed into one of 21 groups upon emergence, with the ten bees in the same group and from the same colony housed together in a wooden hoarding cage (12×12×12 cm). Each group of bees was fed 1 g of pollen mixed with 0.5 mL of syrup (1∶1 sucrose to water by weight), which they fully consumed in 2–4 days. These pollen cakes were placed in small petri dishes with the laboratory cages. Pollen from either one of the crop fields or one of two control diets were used. The pollen control group (“BRL”) was fed a mixed pollen diet prepared by the USDA-ARS Bee Research Laboratory. This pollen was collected in the desert Southwest (Arizona Bee Products, Tucson, AZ) and tested as pesticide-free by the USDA Agricultural Marketing Service prior to use. A protein control group was fed an artificial honey bee pollen substitute, MegaBee®. The Nosema inoculum was freshly prepared by mixing Nosema spores isolated from an infected colony (details provided in [26]) with 50% sucrose solution to obtain a concentration of ca. 2 million spores per 5 mL. We fed 5 mL of the Nosema inoculum to each cage during the first two days of adult life, then provided bees with ad libitum access to clean 50% (w/v) sucrose solution. We collected bees 12 days after infection and examined them for the presence or absence of N. ceranae spores by homogenizing individual abdomens in 1 mL distilled water. Here we focus only on infection prevalence, the number of individuals with Nosema spores.
To look for potential effects of individual pesticides on susceptibility to Nosema infection, we calculated the relative risk and its 95% confidence interval for bees becoming infected after consuming pollen with a specific pesticide. Relative risk measures the chance of developing a disease after a particular exposure [41], here each pesticide. A relative risk value of one indicates that the probability of infection is equal between exposed and non-exposed groups.
We further tested effects of pesticides in pollen on measured Nosema prevalence using a generalized linear mixed model with a bee’s Nosema status as the response variable, the source hive and pesticide variables as fixed effects, and the pollen sample fed to the bee as a random effect. Collinearity prevented developing a full model to investigate in detail how pesticides and pollen source affect bees’ susceptibility to Nosema infection. We thus selected for analysis two measures that vary with crop and are not nested: total pesticide diversity and fungal load. To graph logistic regression results in a meaningful manner, we followed recent recommendations [42], [43] and a modification of the logi.hist.plot function in the R popbio package [44] that shows our mixed model output.
Results
Pollen Collection
Bee colonies collected different amounts of pollen in the different crops (Table 1; Kruskal-Wallis test: H7 = 29.6, p = 0.0001). Pollen diversity, estimated by quantifying the number of differently colored pollen pellets collected in pollen traps, varied by crop (Table 1; Kruskal-Wallis test: H7 = 23.5, p = 0.0014). The proportion of pollen that bees collected from the target crop, except for almond and apple, was low (mean±se = 0.33±0.05; Table S1). Like pollen weights, this proportion dramatically differed between crops (Fig. 1; H7 = 44.86, p<0.0001). Notably, none of the pollen trapped from hives in blueberry, cranberry (early and late), pumpkin or watermelon fields was from the target crop.
thumbnail
Figure 1. Pollen collection from the crop where a hive was located was low for most crops.
Bars show mean ± se. Letters indicate statistically significant differences (p<0.05).
https://doi.org/10.1371/journal.pone.0070182.g001
Pesticide Analysis
All pollen collected in this study contained pesticides (Table 2; mean ± se = 9.1±1.2 different chemicals, range 3–21). Pesticide loads ranged from 23.6 to 51,310.0 ppb (11,760.0±3,734.2 ppb). The maximum pesticide concentration in any single pollen sample exceeded the median lethal dose (LD50, the dose required to kill half a population within 24 or 48 h) for esfenvalerate and phosmet (Table 2). The number of pesticides detected in trapped pollen varied by the crop in which the bee hives were located (Kruskal-Wallis test: H6 = 12.96, p = 0.04), but the total pesticide load did not (H6 = 11.21, p = 0.08)(Fig. 2).
thumbnail
Figure 2. Pesticide diversity found in pollen samples, but not pesticide load, varied by crop.
White bars show pesticide diversity, gray bars show pesticide load (mean ± se). Post-hoc testing found the following groups, where letters indicate statistically significant differences: apple a, b; blueberry c; cranberry_early d; cranberry_late b, d, e, f; cucumber e; pumpkin c, d, f; and watermelon d.
https://doi.org/10.1371/journal.pone.0070182.g002
thumbnail
Table 2. Pesticides found in pollen trapped off honey bees returning to the nest.
https://doi.org/10.1371/journal.pone.0070182.t002
We found insecticides and fungicides in all 19, and herbicides in 23.6% of, pollen samples. Insecticides present in pollen collected by the bees came from seven categories. We found oxadiazines in 10.5%, neonicotinoids in 15.8%, carbamates in 31.6%, cyclodienes in 52.6%, formamidines in 52.6%, organophosphates in 63.2%, and pyrethroids in 100% of pollen samples. Both neonicotinoids and oxadiazines were present only in pollen collected by bees in apple orchards (Figs. 3, S1). Within a sample, pollen fungicide loads were significantly higher than loads of herbicides or any of the insecticide categories (Fig. 4; GLMM, likelihood ratio test: χ2 = 121.9, df = 8, p<0.0001).
thumbnail
Figure 3. Fungicide and neonicotinoid diversities varied by crop.
Bars show the total number of pesticides in each category found in each crop. Kruskal-Wallis test statistics comparing pesticide diversity between crops are: fungicides, H6 = 16.1, p = 0.01; cyclodienes, H6 = 6.9, p = 0.33; neonicotinoids, H6 = 17.9, p = 0.007; organophosphates, H6 = 14.3, p = 0.03; pyrethroids, H6 = 7.8, p = 0.26. We only compared pesticide diversities for categories containing at least three chemicals. Sequential Bonferroni adjusted critical values are: 0.01, 0.0125, 0.0167, 0.025, 0.05. A * indicates that the total number of pesticides varied between crops within that pesticide category.
https://doi.org/10.1371/journal.pone.0070182.g003
thumbnail
Figure 4. Load varied by pesticide category.
Letters indicate statistically significant differences. The total load for each category is weighted by the number of chemicals in that category, to facilitate comparison across categories.
https://doi.org/10.1371/journal.pone.0070182.g004
After adjusting for multiple comparisons, pesticide loads did not vary by crop for any pesticide category (Fig. S1). We calculated pesticide diversity within only those categories containing three or more chemicals. Fungicide and neonicotinoid diversities varied by crop, but diversities of other pesticide categories did not (Fig. 3).
Nosema Infection
147 of the 630 bees (23.3%) fed Nosema spores became infected. 22 of the 35 pesticides (62.9%) found in our pollen samples had relative risk values significantly different from 1 (Table 2). 8 pesticides (22.9%) were associated with increased Nosema prevalence, while the remaining 14 were associated with decreased Nosema prevalence. Two of the three detected pesticides applied by beekeepers to control hive mites (marked with a * in Table 2) had a relative risk larger than two, indicating Nosema prevalence in bees fed pollen containing those chemicals (DMPF and fluvalinate) was more than double the Nosema prevalence in bees that did not consume these chemicals. Of the seven pesticides found in pollen from over half, or at least four, of the crops, the majority were associated with higher Nosema prevalence in bees that consumed them. Both control diets had relative risk values not significantly different from one.
A pollen sample’s fungicide load significantly affected Nosema prevalence among bees fed that pollen (Fig. 5; GLMM, likelihood ratio test: χ2 = 5.8, df = 1, p = 0.02), but pesticide diversity did not (χ2 = 1.7, df = 1, p = 0.19). A bee’s source colony, included as a blocking variable, also did not affect Nosema prevalence (χ2 = 2.0, df = 2, p = 0.36). Replacing fungicide load with chlorothalonil load obtained the same result (chlorothalonil load: χ2 = 5.3, df = 1, p = 0.02; pesticide diversity: χ2 = 1.5, df = 1, p = 0.23; source colony: χ2 = 2.0, df = 2, p = 0.36; fungicide load model AIC = 612.71, chlorothalonil load model AIC = 613.15). Chlorothalonil was also the most abundant fungicide in our samples, and comprised 50.0±10.2% (mean ± se) of the per sample total fungicide load.
thumbnail
Figure 5. Probability of Nosema infection increased with fungicide load in consumed pollen.
Histograms show the number of bees with (top) and without (bottom) Nosema spores as a function of the fungicide load in the pollen they were fed. The curve shows the predicted probability of Nosema infection.
https://doi.org/10.1371/journal.pone.0070182.g005
Discussion
The results from this study highlight several patterns that merit further attention. First, despite being rented to pollinate specific crops, honey bees did not always return to the nest with corbicular pollen from those crops. These findings support other research with honey bees and native bees indicating that in some crops native bees may be more efficient pollinators [45]. Second, fungicides were present at high levels in both crop and non-crop pollen collected by bees. Third, two fungicides (chlorothalonil and pyraclostrobin), and two miticides used by beekeepers to control varroa infestation (amitraz and fluvalinate) had a pronounced effect on bees’ ability to withstand parasite infection. Research on pesticides’ effects on bee health has focused almost exclusively on insecticides (e.g. fipronil [15] and the neonicotinoids imidacloprid [13], [14] and thiacloprid [15]). Finally, several individual pollen samples contained loads higher than the median lethal dose for a specific pesticide. While multiple studies have shown negative effects of specific pesticides on honey bee individual and colony health [14], [15], [22], [26] and high pesticide exposure [27], [28], ours is the first to demonstrate how real world pollen-pesticide blends affect honey bee health.
Our results show that beekeepers need to consider not only pesticide regimens of the fields in which they are placing their bees, but also spray programs near those fields that may contribute to pesticide drift onto weeds. The bees in our study collected pollen from diverse sources, often failing to collect any pollen from the target crop (Fig. 1). All of the non-target pollen that we were able to identify to genus or species was from wildflowers (Table S1), suggesting the honey bees were collecting significant amounts of pollen from weeds surrounding our focal fields. The two exceptions to this were hives placed in almond and apple orchards. Almond flowers early in the year, and almond orchards are large, thus providing honey bees with little access to other flowers. Honey bees rarely collect pollen from blueberry or cranberry flowers, which only release large quantities of pollen after being vibrated by visiting bees (buzz pollination) [46], [47]. Honey bees are not capable of buzz pollination and thus are unlikely to collect large amounts of pollen from these plants to bring back to the colony. Bumble bees, which can buzz pollinate, collect mainly blueberry pollen when placed in blueberry fields [48]. Interestingly, the two crops that saw high levels of pollen collection by honey bees are Old World crops that evolved with honey bees as natural pollinators. Crops native to the New World, where honey bees have been introduced, yielded little or no pollen in our samples.
It is possible that bees were exposed to pesticides while collecting nectar from our focal crops, even when we detected no pollen from those crops. Because pollen traps collect only corbicular pollen intended for consumption by the colony, our data indicate only flowers from which bees are actively collecting pollen and not all flowers they visited. Several studies have detected pesticides in floral nectar and pollen [49], [50], sometimes in concentrations with sublethal effects on honey and bumble bees [51], [52]. Honey bees may collect nectar from blueberry and cranberry flowers via legitimate visits or “robbing” through slits cut at the base of flower corollas [53]. However, exposure to pesticides via nectar may be unlikely in cucumber, pumpkin and watermelon. Beekeepers often report poor honey production when their hives are placed in these crops (pers. obs.).
The combination of high pesticide loads and increased Nosema infection rates in bees that consumed greater quantities of the fungicides chlorothalonil and pyraclostrobin suggest that some fungicides have stronger impacts on bee health than previously thought. Nosema infection was more than twice as likely (relative risk >2) in bees that consumed these fungicides than in bees that did not. Research on the sub-lethal effects of pesticides on honey bees has focused almost entirely on insecticides, especially neonicotinoids [54]. In our study, neonicotinoids entered the nest only via apple pollen. However, we found fungicides at high loads in our sampled crops. While fungicides are typically less lethal to bees than insecticides (see LD50 values in Table 2), these chemicals still have potential for lethal [55] and sub-lethal effects. Indeed, the fungicides chlorothalonil (found at high concentrations in our pollen samples) and myclobutanil increases gut cell mortality to the same degree as imidacloprid [56], an insecticide with numerous sub-lethal effects (e.g. [21], [57]). Exposure to fungicides can also make bees more sensitive to acaricides, reducing medial lethal doses [58]. In our study, consuming pollen with higher fungicide loads increased bees’ susceptibility to Nosema infection. This result is likely driven by chlorothalonil loads. The pesticide with the highest relative risk was the fungicide pyraclostrobin. Bees that consumed pollen containing pyraclostrobin were almost three times as likely (relative risk = 2.85, 95% CI 2.16–3.75; Table 2) than bees consuming pollen without this chemical to become infected after Nosema exposure. Our results show the necessity of testing for sub-lethal effects of pesticides on bees, and advocate for testing more broadly than the insecticides that are the targets of most current research.
A similarly large increased risk of Nosema infection was associated with consumption of DMPF and fluvalinate, miticides applied by beekeepers to help control the highly-destructive Varroa mite [3]. The path from in-hive application of these miticides to pollen on foragers returning to the hive is unclear. An increasingly popular practice, rotating combs out of hives to remove accumulated pesticides, is expected to reduce miticide levels in hives, and will hopefully decrease spread of these chemicals to the environment. Potential extra-nest sources, however, would slow efforts to reduce miticide accumulation and slow the development of resistance to these chemicals.
Insecticide relative risk values showed an interesting pattern: directional separation by insecticide family. Within a family, relative risk values significantly different than one were almost all in the same direction. The formamidine (DMPF) and two of the three the pyrethroids (bifenthrin and fluvalinate, but not esfenvalerate) were associated with an increased risk of Nosema infection. The carbamate (carbaryl), all neonicotinoids (acetamiprid, imidacloprid and thiacloprid), organophosphates (coumaphos, diazinon and phosmet) and the oxadiazine (indoxacarb) were associated with reduced risk of Nosema infection. Esfenvalerate and coumaphos have previously been found to be associated with apiaries without Colony Collapse Disorder [59]. These patterns suggest that insecticides’ modes of action have differential effects on honey bee immune functioning. Because of the relatively small number of pesticides we found in each insecticide family, however, additional sampling is necessary to determine how robust this pattern is.
The large numbers of pesticides found per sample and the high concentrations of some pesticides are concerning. First, two pollen samples contained one pesticide each at a concentration higher than the median lethal dose. Esfenvalerate (LD50 = 0.13 ppm) was measured at 0.216 ppm in pollen collected by bees in a cucumber field, and phosmet (LD50 = 8.83 ppm) at 14.7 ppm in one apple orchard. While the mean loads for these pesticides are well below their respective median lethal doses (0.0169 ppm for esfenvalerate, 0.7987 ppm for phosmet), our data indicate some bee colonies are being exposed to incredibly high levels of these chemicals. Second, research suggests that simultaneous exposure to multiple pesticides decreases lethal doses [58], [60] or increases supersedure (queen replacement) rate [61]. Our pollen samples contained an average of nine different pesticides, ranging as high as 21 pesticides in one cranberry field. Thus published LD50 values may not accurately indicate pesticide toxicity inside a hive containing large numbers of pesticides. Research looking at additive and synergistic effects between multiple pesticides is clearly needed. Third, pesticides can have sub-lethal effects on development, reproduction, learning and memory, and foraging behavior. The mean and maximum imidacloprid loads in our samples (0.0028 and 0.0365 ppm, respectively) are higher than some published imidacloprid concentrations with sub-lethal effects on honey and bumble bees (0.001–0.0098 ppm [21], [54], [62]).
It is not surprising that total pollen collection varied by crop. Bee foraging activity levels vary with weather [63], thus outcomes of short-term measurements may be sensitive to temperature, cloud cover or humidity during data collection. Because we collected pollen samples from different parts of the country and on different days, weather conditions undoubtedly differed between crops. Crop flowering timing and landscape-level floral availability can also affect bee activity levels. We focused our analyses on variables less affected by these factors, such as the diversity of pollen types found in samples and the proportion of a sample that was from the target crop.
Our results are consistent with previously published pesticide analyses of pollen collected by honey bees or honey bee nest material [16], [18], [27]. The more intensive and geographically more diverse sampling of Mullin et al. [27] resulted in almost triple the number of pesticides we found, but the average number of pesticides per sample (7.1) is slightly lower than our 9.1. In our study and those listed above, pesticides applied by beekeepers to control hive pests were present in a large proportion of the samples, often in quantities higher than most of the pesticides that are applied to crops.
Our results combined with several recent studies of specific pesticides’ effects on Nosema infection dynamics [13]–[15] indicate that a detrimental interaction occurs when honey bees are exposed to both pesticides and Nosema. Specific results vary, and may depend on the pesticide or dose used. For example, bees exposed to imidacloprid and Nosema can have lower spore counts than bees only infected with the pathogen but also exhibit hindered immune functioning [13]. Our study improves on previous methodologies by feeding pollen with real-world pesticide blends and levels that truly represents the types of exposure expected with pollination of agricultural crops. The significant increase in Nosema infection following exposure to the fungicides in pollen we found therefore indicates a pressing need for further research on lethal and sub-lethal effects of fungicides on bees. Given the diverse routes of exposure to pesticides we show, and increasing evidence that pesticide blends harm bees [16], , there is a pressing need for further research on the mechanisms underlying pesticide-pesticide and pesticide-disease synergistic effects on honey bee health.
Supporting Information
Figure S1.
Pesticide loads did not differ by crop for any pesticide category. Kruskal-Wallis test statistics comparing pesticide loads between crops are: fungicides, H6 = 10.6, p = 0.10; herbicides, H6 = 8.3, p = 0.22; carbamates, H6 = 13.4, p = 0.04; cyclodienes, H6 = 6.7, p = 0.35; formamidines, H6 = 13.6, p = 0.03; neonicotinoids, H6 = 17.8, p = 0.007; organophosphates, H6 = 14.5, p = 0.02; oxadiazines, H6 = 11.3, p = 0.08; pyrethroids, H6 = 9.6, p = 0.14. Sequential Bonferroni adjusted critical values are: 0.0055, 0.0063, 0.0071, 0.0083, 0.01, 0.0125, 0.0167, 0.025, 0.06.
https://doi.org/10.1371/journal.pone.0070182.s001
(DOCX)
Table S1.
Plant sources of pollens collected by bees placed in seven crops.
https://doi.org/10.1371/journal.pone.0070182.s002
(DOCX)
Acknowledgments
We thank David Hackenberg and David Mendes for letting us work with their bee hives, John Baker and Rob Snyder for field assistance, Roger Simonds for pesticide identification, and Vic Levi and Nathan Rice for assistance with Nosema assays.
The views expressed in this article are those of the authors and do not necessarily represent the policies or positions of the US Department of Agriculture (USDA).
Author Contributions
Conceived and designed the experiments: JSP RR DV. Performed the experiments: JSP MA JS DV. Analyzed the data: EML DV. Wrote the paper: JP EML DV.
References
1. Klein A-M, Vaissiere BE, Cane JH, Steffan-Dewenter I, Cunningham SA, et al. (2007) Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society B-Biological Sciences 274: 303–313.
2. vanEngelsdorp D, Meixner MD (2010) A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. Journal of Invertebrate Pathology 103: S80–S95.
3. vanEngelsdorp D, Caron D, Hayes J, Underwood R, Henson M, et al. (2012) A national survey of managed honey bee 2010–11 winter colony losses in the USA: results from the Bee Informed Partnership. Journal of Apicultural Research 51: 115–124.
4. van der Zee R, Pisa L, Andonov S, Brodschneider R, Charriere JD, et al. (2012) Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008–9 and 2009–10. Journal of Apicultural Research 51: 91–114.
5. Aizen MA, Garibaldi LA, Cunningham SA, Klein AM (2009) How much does agriculture depend on pollinators? Lessons from long-term trends in crop production. Annals of Botany 103: 1579–1588.
6. Gallai N, Salles J-M, Settele J, Vaissiere BE (2009) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecological Economics 68: 810–821.
7. NRC (2007) Status of pollinators in North America. Washington, DC: The National Academies Press. 312 p.
8. Biesmeijer JC, Roberts SPM, Reemer M, Ohlemuller R, Edwards M, et al. (2006) Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313: 351–354.
9. vanEngelsdorp D, Speybroeck N, Evans JD, Nguyen BK, Mullin C, et al. (2010) Weighing risk factors associated with bee colony collapse disorder by classification and regression tree analysis. Journal of Economic Entomology 103: 1517–1523.
10. Cornman RS, Tarpy DR, Chen Y, Jeffreys L, Lopez D, et al. (2012) Pathogen webs in collapsing honey bee colonies. PloS ONE 7: e43562.
11. Foley K, Fazio G, Jensen AB, Hughes WOH (2012) Nutritional limitation and resistance to opportunistic Aspergillus parasites in honey bee larvae. Journal of Invertebrate Pathology 111: 68–73.
12. Alaux C, Ducloz F, Crauser D, Le Conte Y (2010) Diet effects on honeybee immunocompetence. Biology Letters 6: 562–565.
13. Alaux C, Brunet J-L, Dussaubat C, Mondet F, Tchamitchan S, et al. (2010) Interactions between Nosema microspores and a neonicotinoid weaken honeybees (Apis mellifera). Environmental Microbiology 12: 774–782.
14. Pettis JS, vanEngelsdorp D, Johnson J, Dively G (2012) Pesticide exposure in honey bees results in increased levels of the gut pathogen Nosema. Naturwissenschaften 99: 153–158.
15. Vidau C, Diogon M, Aufauvre J, Fontbonne R, Vigues B, et al. (2011) Exposure to sublethal doses of fipronil and thiacloprid highly increases mortality of honeybees previously infected by Nosema ceranae. PLoS ONE 6: e21550.
16. Wu JY, Smart MD, Anelli CM, Sheppard WS (2012) Honey bees (Apis mellifera) reared in brood combs containing high levels of pesticide residues exhibit increased susceptibility to Nosema (Microsporidia) infection. Journal of Invertebrate Pathology 109: 326–329.
17. Desneux N, Decourtye A, Delpuech J-M (2007) The sublethal effects of pesticides on beneficial arthropods. Annual Review of Entomology 52: 81–106.
18. Wu JY, Anelli CM, Sheppard WS (2011) Sub-lethal effects of pesticide residues in brood comb on worker honey bee (Apis mellifera) development and longevity. PLoS ONE 6: e14720.
19. Eiri D, Nieh JC (2012) A nicotinic acetylcholine receptor agonist affects honey bee sucrose responsiveness and decreases waggle dancing. Journal of Experimental Biology 215: 2022–2029.
20. Gregorc A, Evans JD, Scharf M, Ellis JD (2012) Gene expression in honey bee (Apis mellifera) larvae exposed to pesticides and Varroa mites (Varroa destructor). Journal of Insect Physiology 58: 1042–1049.
21. Whitehorn PR, O'Connor S, Wackers FL, Goulson D (2012) Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 336: 351–352.
22. Boncristiani H, Underwood R, Schwarz R, Evans JD, Pettis J, et al. (2012) Direct effect of acaricides on pathogen loads and gene expression levels in honey bees Apis mellifera. Journal of Insect Physiology 58: 613–620.
23. Cameron SA, Lozier JD, Strange JP, Koch JB, Cordes N, et al. (2011) Patterns of widespread decline in North American bumble bees. Proceedings of the National Academy of Sciences 108: 662–667.
24. George PJE, Ambrose DP (2004) Impact of insecticides on the haemogram of Rhynocoris kumarii Ambrose and Livingstone (Hem., Reduviidae). Journal of Applied Entomology 128: 600–604.
25. Delpuech JM, Frey F, Carton Y (1996) Action of insecticides on the cellular immune reaction of Drosophila melanogaster against the parasitoid Leptopilina boulardi. Environmental Toxicology and Chemistry 15: 2267–2271.
26. Chaimanee V, Chantawannakul P, Chen Y, Evans JD, Pettis JS (2012) Differential expression of immune genes of adult honey bee (Apis mellifera) after inoculated by Nosema ceranae. Journal of Insect Physiology 58: 1090–1095.
27. Mullin CA, Frazier M, Frazier JL, Ashcraft S, Simonds R, et al. (2010) High levels of miticides and agrochemicals in North American apiaries: implications for honey bee health. PLoS ONE 5: e9754.
28. Chauzat MP, Faucon JP, Martel AC, Lachaize J, Cougoule N, et al. (2006) A survey of pesticide residues in pollen loads collected by honey bees in France. Journal of Economic Entomology 99: 253–262.
29. Johnson RM, Wen Z, Schuler MA, Berenbaum MR (2006) Mediation of pyrethroid insecticide toxicity to honey bees (Hymenoptera : Apidae) by cytochrome P450 monooxygenases. Journal of Economic Entomology 99: 1046–1050.
30. Higes M, Martin-Hernandez R, Botias C, Garrido Bailon E, Gonzalez-Porto AV, et al. (2008) How natural infection by Nosema ceranae causes honeybee colony collapse. Environmental Microbiology 10: 2659–2669.
31. Higes M, Martin-Hernandez R, Meana A (2010) Nosema ceranae in Europe: an emergent type C nosemosis. Apidologie 41: 375–392.
32. Farrar CL (1947) Nosema losses in package bees as related to queen supersedure and honey yields. Journal of Economic Entomology 40: 333–338.
33. Konietschke F (2011) nparcomp v. 1.0–1. R package.
34. Crompton CW, Wojtas WA (1993) Pollen grains of Canadian honey plants. Ottawa: Agriculture Canada. 228 p.
35. Hodges D (1964) Pollen grain drawings from the pollen loads of the honeybee. London: Bee Research Association. 51 p.
36. Kirk WDJ (2006) A colour guide to pollen loads of the honey bee. Cardiff, UK: International Bee Research Association. 54 p.
37. Kearns CA, Inouye DW (1993) Techniques for Pollination Biologists. Niwot, CO: University Press of Colorado. 583 p.
38. Wilson EE, Sidhu CS, LeVan KE, Holway DA (2010) Pollen foraging behaviour of solitary Hawaiian bees revealed through molecular pollen analysis. Molecular Ecology 19: 4823–4829.
39. Cane JH, Snipes S (2006) Characterizing floral specialization by bees: analytical methods and a revised lexicon for oligolecty. In: Waser NM, Ollerton J, editors. Plant-Pollinator Interactions: From Specialization to Generalization. Chicago: University of Chicago Press. 99–122.
40. Holm S (1979) A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics 6: 65–70.
41. vanEngelsdorp D, Lengerich E, Spleen A, Dainat B, Cresswell J, et al. (2013) Standard epidemiological methods to understand and improve Apis mellifera health. The COLOSS BEEBOOK: Volume II: Standard methods for Apis mellifera pest and pathogen research. Journal of Apicultural Research 52: 1–16.
42. de la Cruz Rot M (2005) Improving the presentation of results of logistic regression with R. Bulletin of the Ecological Society of America. 86: 41–48.
43. Smart J, Sutherland WJ, Watkinson AR, Gill JA (2004) A new means of presenting the results of logistic regression. Bulletin of the Ecological Society of America 85: 100–102.
44. Stubben C, Milligan B, Nantel P (2012) popbio: Construction and analysis of matrix population models v. 2.4. R package.
45. Garibaldi LA, Steffan-Dewenter I, Winfree R, Aizen MA, Bommarco R, et al. (2013) Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 339: 1608–1611.
46. Delaplane KS, Mayer DF (2000) Crop Pollination by Bees. New York: CABI Publishing. 344 p.
47. Kremen C (2008) Crop pollination services from wild bees. In: James RR, Pitts-Singer TL, editors. Bee Pollination in Agricultural Ecosystems. New York: Oxford University Press. 10–26.
48. Whidden TL (1996) The fidelity of commercially reared colonies of Bombus impatiens Cresson (Hymenoptera: Apidae) to lowbush blueberry in southern New Brunswick. The Canadian Entomologist 128: 957–958.
49. Choudhary A, Sharma DC (2008) Dynamics of pesticide residues in nectar and pollen of mustard (Brassica juncea (L.) Czern.) grown in Himachal Pradesh (India). Environmental Monitoring and Assessment 144: 143–150.
50. Krischik VA, Landmark AL, Heimpel GE (2007) Soil-applied imidacloprid is translocated to nectar and kills nectar-feeding Anagyrus pseudococci (Girault) (Hymenoptera : Encyrtidae). Environmental Entomology 36: 1238–1245.
51. Stoner KA, Eitzer BD (2012) Movement of soil-applied imidacloprid and thiamethoxam into nectar and pollen of squash (Cucurbita pepo). PLoS ONE 7: e39114.
52. Dively GP, Kamel A (2012) Insecticide residues in pollen and nectar of a cucurbit crop and their potential exposure to pollinators. Journal of Agricultural and Food Chemistry 60: 4449–4456.
53. Sampson BJ, Danka RG, Stringer SJ (2004) Nectar robbery by bees Xylocopa virginica and Apis mellifera contributes to the pollination of rabbiteye blueberry. Journal of Economic Entomology 97: 735–740.
54. Blacquiere T, Smagghe G, van Gestel CAM, Mommaerts V (2012) Neonicotinoids in bees: a review on concentrations, side-effects and risk assessment. Ecotoxicology 21: 973–992.
55. Hooven LA (2013) Effect of fungicides on development and behavior of honey bees. American Bee Research Conference. Hershey, PA.
56. Gregorc A, Ellis JD (2011) Cell death localization in situ in laboratory reared honey bee (Apis mellifera L.) larvae treated with pesticides. Pesticide Biochemistry and Physiology 99: 200–207.
57. Gill RJ, Ramos-Rodriguez O, Raine NE (2012) Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature 491: 105–108.
58. Johnson RM, Dahlgren L, Siegfried BD, Ellis MD (2013) Acaricide, fungicide and drug interactions in honey bees (Apis mellifera). PLoS ONE 8: e54092.
59. vanEngelsdorp D, Evans JD, Saegerman C, Mullin C, Haubruge E, et al. (2009) Colony Collapse Disorder: a descriptive study. PLoS ONE 4: e6481.
60. Iwasa T, Motoyama N, Ambrose JT, Roe RM (2004) Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera. Crop Protection 23: 371–378.
61. Drummond FA, Aronstein K, Chen YP, Eitzer B, Ellis J, et al.. (2013) Colony losses in stationary apiaries related to pesticide contamination of pollen. American Bee Research Conference. Hershey, PA.
62. Laycock I, Lenthall KM, Barratt AT, Cresswell JE (2012) Effects of imidacloprid, a neonicotinoid pesticide, on reproduction in worker bumble bees (Bombus terrestris). Ecotoxicology 21: 1937–1945.
63. Danka RG, Sylvester HA, Boykin D (2006) Environmental influences on flight activity of USDA-ARS Russian and Italian stocks of honey bees (Hymenoptera : Apidae) during almond pollination. Journal of Economic Entomology 99: 1565–1570.
64. US EPA (1997) Fact Sheet for Azoxystrobin. Washington, DC: US EPA. 23 p.
65. US EPA (1999) Reregistration Eligibility Decision (RED) for Captan. Washington, DC: US EPA. 250 p.
66. US EPA (1999) Reregistration Eligibility Decision (RED) for Chlorothalonil. Washington, DC: US EPA. 337 p.
67. US EPA (1998) Pesticide Fact Sheet for Cyprodinil. Washington, DC: US EPA. 13 p.
68. OPP Pesticide Toxicity Database: Ecotox Results for Difenoconazole website. Available: http://www.ipmcenters.org/Ecotox/Detail ... rdID=10104. Accessed 2013 Jan 15.
69. Atkins EL (1988) RH-7592 technical: bee adult toxicity dusting test. Spring House, PA: Rohm and Haas Company. 6 p.
70. ECCO-Team (2012) Full Report on Pyraclostrobin. York, UK: Pesticides Safety Directorate. 208 p.
71. US EPA (2006) Reregistration Eligibility Decision for Pentachloronitrobenzene. Washington, DC: US EPA. 127 p.
72. Carleton J (1999) Carfentrazone-ethyl Herbicide Environmental Fate and Ecological Effects Assessment and Risk Characterization for a Section 3 for Use on Sweet Corn, Sorghum, and Rice. Washington, DC: US EPA. 25 p.
73. US EPA (1997) Reregistration Eligibility Decision (RED) for Pendimethalin. Washington, DC: US EPA. 239 p.
74. Atkins EL, Kellum D (1981) Effect of pesticides on agriculture: maximizing the effectiveness of honey bees as pollinators. 1981 Report of research to California Alfalfa Seed Production Research Board. Philadelphia, PA: FMC Corporation. 2 p.
75. US EPA (2004) Interim Reregistration Eligibility Decision (IRED) for Carbaryl. Washington, DC: US EPA. 305 p.
76. Cox C (1994) Insecticide fact sheet: cyfluthrin. Journal of Pesticide Reform 14: 28–34.
77. Gough HJ, Collins IG, Everett CJ, Wilkinson W (1984) PP321: acute contact and oral toxicity to honey bees (Apis mellifera). Wilmington, DE: ICI Americas Inc. 7 p.
78. US EPA (2008) Reregistration Eligibility Decision (RED) for Cypermethrin. Washington, DC: US EPA. 113 p.
79. Pesticide Management Education Program: Pesticide Information Profile: Deltamethrin website. Available: http://pmep.cce.cornell.edu/profiles/ex ... n-ext.html. Accessed 2013 Jan 15.
80. US EPA (2004) Interim Reregistration Eligibility Decision (IRED) for Diazinon. Washington, DC: US EPA. 134 p.
81. Hoxter KA, Smith GJ (1990) H-#18, 151 (Technical Asana): an acute contact toxicity study with the honey bee. Wildlife International Ltd. 4 p.
82. US EPA (2005) Reregistration Eligibility Decision (RED) for Tau-fluvalinate. Washington, DC: US EPA. 85 p.
83. Decourtye A, Lacassie E, Pham-Delegue MH (2003) Learning performances of honeybees (Apis mellifera L) are differentially affected by imidacloprid according to the season. Pest Management Science 59: 269–278.
84. Abel S (2005) Environmental fate & effects division risk assessment for proposed new users of indoxacarb on grapes, fire ants, mole crickets, alfalfa, peanuts, soybeans. Washington, DC: US EPA. 263 p.
85. Johansen CA, Mayer DF (1990) Pollinator Protection: A Bee & Pesticide Handbook. Cheshire, CN: Wicwas Press. 212 p.
86. US EPA (1998) Reregistration Eligibility Decision (RED) for Methomyl. Washington, DC: US EPA. 266 p.
87. US EPA (1992) R.E.D. Facts for Heptachlor. Washington, DC: US EPA. 4 p
http://journals.plos.org/plosone/articl ... ne.0070182
Users browsing this forum: No registered users and 175 guests